In a recent paper entitled "Paleoelevation in the Jianchuan Basin of the southeastern Tibetan Plateau based on stable isotope and pollen grain analyses", Wu, Zhang et al. find that "... pollen data from the Shuanghe Fm indicates a paleoelevation of 1.3–2.6 km above sea level (asl), which would most probably have been associated with a vegetation cover consisting of tropical [most taxa in the list are however extra-tropical]-subtropical, deciduous, coniferous, broadleaf forests. The reconstructed mean annual air temperature (MAAT) had a value of 16.8–21.7 °C, warmer than today's MAAT (~6 °C)." A result in some match with their stable isotope of the Jiuziyan Fm. "... suggest[ing] that the surface of the Jianchuan Basin was at a paleoelevation between 0.5+0.8–0.5 km asl (i.e. sea-level) and 2.5 ± 0.7 km asl (δ18Omw: −8.9 ± 1.3‰, 2σ)"
You can never be wrong with low to mid-altitude subtropical in the Eocene, can you?
Only that the fossil flora is composed mainly of conifers and other meso- to microthermic elements (as stated by the authors on p. 9), which would be very atypical for a tropical-subtropical setting, but typical for a temperate-boreal. But as we showed (still ignored but also not rebutted by Palaeo^3 authors and editors), all roads lead to Yunnan's subtropical climate when using Chinese palaeofloras (Grimm et al. 2016).
 |
| Distribution of Pinus (source: Wikimedia Commons). Clearly a genus indicative for mild temperate to tropical conditions (Tann > 7 according Wu et al., table 1) |
The higher we go up, the colder it gets
As most of us know (but not users of the Coexistence Approach or editors/peers of PPP, as we will see). With dropping temperatures, the likelihood for snow increases, so you see different forests/ vegetation belts when roaming mountaineous regions. When you go to southern Hokkaido with its Dfb climate (snow but proper summers), you'll find beech forests (Fagus crenata, the sister species of the western Eurasian beech, F. sylvatica), at sea-level; same in southern Canada (F. grandifolia, a distant relative of both) or southern Scandinavia. Going to Honshu or even Taiwan, one can readily notice that the beech forest belt goes up the mountain: a thumb rule is that it starts at about 500 m in northern Honshu (more or less fitting the border between Cfa and Cfb climate), increasing to 1000/1500 m in the subtropical, southern Honshu, Ryukyu or near-tropical (at sea-level) Taiwan.
 |
| Altitudinal and latitudinal range of beech in Japan (see Maycock 1994, compiled for Grimm & Denk 2012, ES6) |
[Some more bioclimatic trivia for people doing quantitative climate reconstruction using plant taxon lists: Being one of the most-constrained (ecology and climate-wise), non-relict tree genera, the genus Fagus has a Cfa, Cfb, Dfb 'Köppen signature' (Denk et al. 2013), meaning it can occur in these three climates, altitudinal or latitudinal. And this seems to have been the case since its very origin. If you have beeches, you can be pretty sure the local climate has been a warm temperate (not to be confused with temperate), mild snow climate with ample precipitation throughout the year (beeches, in contrast to many other 'nemoral' elements, do not occur in Cwa or Cwb climates; not even in China). Nevertheless, the hottest beeches can be found in northern Florida close to nice summer retreats (and beaches) at Tann of 19–20 °C (Maycock 1994, still subtropical). In Georgia (the country, not the U.S. state), the beech goes down into the subtropical zone and to sea-level (Tann ~ 18 °C), as well as some Chinese species in China's Cfa climate zone. But it doesn't tolerate Dfc climates, too short summers, in contrast to some species of its even more common, distant relative, the oaks; the coldest North American beech stands have nonetheless a Tann of ~4 °C. This is the reason the Coexistence Approach does need relicts with unrepresentative, much too narrow modern climate intervals. Otherwise, the result would always be something between 5–15 °C MA(A)T (temperate forests) and 15°–25 °C (tropical), and not the much wanted 15.6°–15.7° C ... For those interested in beech-climate correlation, info has been compiled in File ES6 to Grimm & Denk 2012.]
In many mountain areas the decrease in temperature the higher we go up approaches the so-called moist-adiabatic lapse rate: On average, we lose 1 °C for every 200 m we go up. Which is also the case for e.g. the vast Himalayan mountain range (especially its southern and eastern slopes) that was the focus of Wu et al.'s study. As seen in the following graph.
 |
| A = Qamdo, Tibet (border between temperate Cwb and Dwb climates; WMO station nr. 5613700); B = Lancang, Yunnan (WMO 5695400; subtropical Cwa) and C = Baingoin, Tibet (arctic-alpine ET; WMO 5527900); note the good fit with the moist-adiabatic lapse rate of 1 °C/200 m using Qamdo as reference point (station data from Lieth et al. 1999). |
Reconstructing palaeo-altitude and -temperature using the Coexistence Approach
How can overlap in the modern altitudinal range (in Tibet) of botanical composite taxa (genera, families), most of which are found across modern China and are covering anything between near-tropical to boreal climates (according to the tolerances reported by Wu et al.), inform about a palaeo-altitude of a "tropical-subtropical mixed deciduous, coniferous broadleaf forest before ~36 Ma" and, independently, the local temperature?
Wu et al. show how this works in their figure 4; the tolerance ranges per taxon are provided in their table 1 — a documentation usually missing in many papers using the method, and nearly all papers published by the NECLIME group, the German-centred (and -funded) Coexistence Approach syndicate.
 |
| Copied from Wu et al. (2018, p. 6). |
The ancient cedars also define the upper boundary of the (palaeo-)temperature interval ("MAAT" ≡ Tann < 21.7 °C), whereas Betula – the birch, one of the most frost- and cold-resistant broadleaved genera of the Northern Hemisphere – and Tsuga – a temperate conifer found today only in eastern North America and East Asia; a classic disjunct distribution for a relict that has seen better times – define the lower boundary for a (warm-)subtropical Tann (> 16.8 °C).
For the altitude "coexistence interval", many NLRs were apparently removed/ignored: Podocarpus, Lygodium, Meliaceae, Carya, Castanea (the chestnut) as too low, and Abies (firs), Picea (spruces), Chenopodiaceae, Ephedra (an odd plant genus, with a near-cosmopolitan distribution, but apparently not in Tibet...), Artemesia (ditto, but common and not odd) as too high. The authors do not provide any clues or guidelines for their elimination strategy.
It may have been informed by the isotope data results.
 |
| Wu et al.'s Fig. 9. May also isotope analysis rest in peace. Note the blue and red doodle lines. |
First alarm-flag missed by peers (?) and unwitting editors: NLR temperature tolerances overlap but not those of the altitudes
Following the (non-sensical) Coexistence Approach protocol for eliminating outliers, there would have been two alternative "statistically supported" (using CA-language, not to be confused with actual statistics) palaeo-altitude coexistence intervals: 1300–1600 m and 2000–2100 m, both with a maximum of 20 out of 25 coexisting. A third equally "supported" interval would be 1700–1700 m — no mistyper here, just following the example of actual CA-papers purporting occasionally MA(A)T CA-intervals such as 15.6–15.6 °C (see Grimm & Denk 2012, ES 2, included in the freely accessible online supporting archive).
But 'outliers' cannot be simply removed. If the ranges of all NLRs don't overlap ("100% coexistence") for a fossil assemblage, then there must have been a shift in one or many lineages.
 |
| Fig. 1 Niche shift over time, one of the reasons for not having 100% overlap when using NLR tolerances for a fossil flora: the "coexistence interval" of the fossils differs from that of the modern counterparts (the "nearest-living relatives"); mutual (climate) range approaches such as the Coexistence Approach assume no shift. Other reasons for missing overlap include mixing across geographic or altitudinal gradients and unrepresentative tolerances (both likely applies to Wu et al.'s reconstruction). |
Here the actuo-palaeontological principle, the very basis for the Coexistence Approach, means:
- The fossil members of the lineages represented by their NLRs had the same temperature niche than today, which is doubtful for not a few members.
- The fossil members of the lineages occurred at the same altitude in the focal area than today, which is effectively untestable.
And the altitude interval must lie within the temperature tolerance (just using common sense, although we were once informed by an "anonymous" (likely Torsten Utescher, the High Wizard of NECLIME, see below), well-hung (it took the peer more than half a year to complete) review of our 2012 paper [PDF including our responses] that especially in the mountains of China, plants occur today outside their natural niche, hence, our critical comparisons were unfair).
Thanks to confidential peer (or pal) review, such logical and practical flaws have never been an issue (a recent example) for the NECLIME syndicate and papers reviewed by a member of the syndicate.
The data the Coexistence Approach works with
The Coexistence Approach is an unweighted mutual (climate) range technique that just uses min- and max-tolerances. This is naturally very crude and has many problems even for reconstructing obvious shifts in the most recent past (Thompson et al. 2012), and even more when being combined with the actua-palaeontological principle (Grimm & Potts 2016). Such as pseudo-coexistence:
 |
| Pseudo-coexistence, the basics — although the (bivariate) climate niches ('envelopes') of the two species don't overlap, hence, you won't find them today growing together, they still produce to finely resolved (narrow) CA intervals. |
 |
| Pseudo-coexistence when mixing floras (e.g. pollen from different altitudes). There are two more or less broad real mutual climate ranges shared by each community, and the completely artificial CA interval (something we Germans call: kleinster gemeinsamer Nenner). |
To understand how min and max of often not independent (again see Thompson et al. 2012) parameters relate (or don't), x-y plots like the ones above establishing/showing the bivariate parameter (climate) space (see e.g. Thompson et al. 1999–2012) can be very enlightening. In Wu et al.'s case it should be very clean, since altitude and temperate are strictly correlated expressed by the moist-adiabatic lapse rate.

Now, let's plot the real envelopes defined by the min-max tolerance ranges for those NLRs where Wu et al. provide both tolerances. First the ones indicative for an altitude (in the focus region) lower than 2500 m...

... and the ones found only at altitudes > 1000 m:

In the case of the low-landers, their elevation tolerance fits their temperature tolerance: the climate envelopes, boxes, are higher than wide. But for those preferring higher altitudes, i.e. those apparently sensitive to too high temperature (subtropical climate), one, the CA-interval determining Cedrus, is does occur at altitudes in the Himalayas outside their temperature tolerance (> 2000 m), an example for High Wizard Utescher's observation that some trees occur today too high up in the Chinese mountains. Ephedra is also climbing too high up; according to its temperate tolerance it should not be able to go beyond 4200 m. It also misses out quite some opportunities at lower elevations since its temperature tolerance would allow growing at sea-level. The reason for this may simply be the observed species / species group, which doesn't behave like others in its genus. Including those that may have lived in the area 35 myrs ago.
There are naturally more examples where Wu et al.'s tolerances are already at odds with each other (not to mention reality). Here's the full plot of their NLRs (which – by the way – should be obligatory to show in any study aiming to do similar things).

Quite a mess. As usual, why you never find bivariate plots in Coexistence Approach papers (but check out the bioclimate atlases for North America by Thompson et al. 1999–2012).
Essentially we can depict four main groups "informing" the palaeo-parameter reconstruction:
- low- to mid-altitude NLRs (green rectangles, see above) with no or differing preference regarding temperature;
- temperature-restricted NLRs (blue rectangles, see below) with no particular preference regarding altitude — occurring at altitudes outside their used temperature tolerances (most-extreme: Betula, birch, used with a 5 °C temperature tolerance but a 4000 m range ≅ 20 °C!), and including all CA-informative NLRs, and
- mid- to high-altitude NLRs (red rectangles, see above) with completely uninformative temperature ranges (from Tann < 0, polar-alpine climates to Tann > 25, tropical, lowland climates, e.g. Abies, spruces).
- The rest are NLRs (yellow rectangles, below) that can occur in any climate (according to the used temperature interval) and any height (sea-level to high-montane).
 |
| Not making sense at all — the blue group, temperature-sensitive, but not in Tibet it seems. |
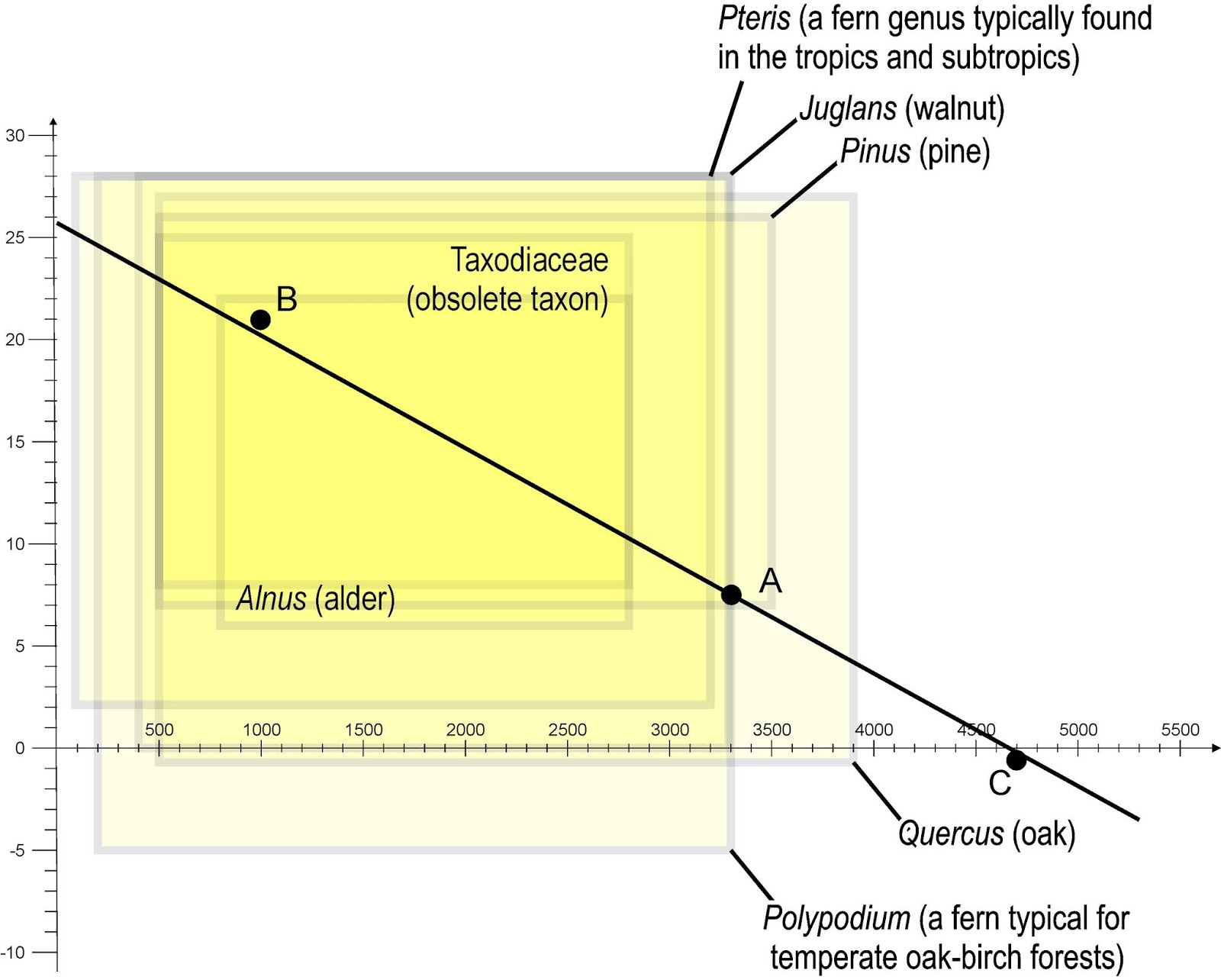 |
| The filling of all CA studies — largely uninformative NLRs. Note that Pteris, according to e.g. Flora of China a tropical-subtropical species-rich fern genus, is recorded with pretty much the same tolerances than Polypodium, a fern with ~ 10 species found "throughout the N[orth] temperate zone" (again: Flora of China, maybe worth a read for CA-applicants working on Chinese palaeofloras) |
Standard when using the Coexistence Approach: ignoring the actuo-palaeontological principle
When the general situation was exactly the same as today and the actuo-palaeontological principle applies (largely), the temperature interval would directly inform the altitude, or vice versa.
Wu et al. argue that in the Eocene the plants grew in a climate with a Tann of c. 6 °C more (see their Conclusion) than today in the same area "... the present climate, which has a MAAT of 13.5 ± 1.4 °C (http://data.cma.cn/site/index.html [try the link]), is much cooler than that of the Eocene." and note the fit with the corresponding global situation.
They further propose "Based on the climatic preferences of the NLRs ... the existence of a warm, subtropical climate, with a MAAT of 16.8–21.7 °C, and a mean value of 19.3 ± 2.5 °C." Not Tibet anymore, but modern-day Yunnan (see Grimm et al. 2016), at about 1000–1500 m (a Cwa climate with very rich mixed evergreen-deciduous subtropical forests and some accessory conifers).
With "mean value" Wu et al. refer to the "center value", which has, so far, no theoretical or practical basis and is defined by the (arithmetic) mean of the upper and lower boundary of the inferred coexistence interval. The mutually shared climate range, Coexistence Approach-based or not, is literally a range: 16.8–21.7 °C would indicate that the temperature was 3° to 8 °C higher than today, and not 5.5 °C ± 2.5 °C higher.
 |
| Close-up on Yunnan and Tibet, showing climate stations included in Lieth et al. (1999). The Cwa station Lincang, Lancang, Simao and Mengzi have Tann within Wu et al.'s coexistence interval, Lijing matches their modern-day reference (Tann = 13.4 °C). Indicated are also Qamdo (Cwc) and Baingoin (ET) used before. |
But in a historical context, there is a slight complication: what if the temperature was raised in general, i.e. globally? Then it may also had been raised locally. A plant affected by this temperature change will evade to or can expand into higher altitudes (and latitudes). If temperatures went down, macrothermic and mesothermic elements would have to retreat to lower altitudes, while microthermic expanded. Something not entirely new to science, but long known for e.g. the beech (Fagus) during Plio-/Pleistocene temperature fluctuations and the subtropical-temperate Nothofagaceae in the Southern Hemisphere (e.g. on Papua New Guinea, in the tropics, Nothofagaceae forests are today starting at c. 1500 m a.s.l. (transition between tropical and subtropical zone) but were down to ~300 m during the glacial maxima. If the NLR occurs at e.g. 1500–2500 m a.s.l. today at 10–15 °C, and the temperature was 10 °C higher in the past in the area, an ancient temperature-sensitive member of the same lineage could only have thrived at 3500–4500 m
Let's take Wu et al. data for granted (which I would not recommend) and assume that the critical plants didn't change their temperature niche in the last 35 Ma (i.e. the temperature tolerance fulfil the actuo-palaeontological principle). The is little reason to assume that the moist-adiabatic lapse rate didn't apply back then (the geotectonic situation is not that different from today). Thus, they would have had to grow higher up than their modern counterparts, i.e. not at 900–1600 m a.sl. (the altitude at which we have today 17–22 °C) or 1300–1600 m a.s.l. (the actual CA-result) but at 1400–2200 m (min. scenario) or 2400–3100 m (max. scenario).
If they would have grown at the same height than their modern (Tibetan) counterparts (i.e. altitude tolerance fulfil the actuo-palaeontological principle), some of their temperature tolerances and preferences must have been higher than today, because the temperature at 1300–1600 m was 20–21.5 °C (yearly average) for the min. scenario (+3 °C) or 25–26.5 °C (+8 °C), i.e. (near)tropical, in the not only regionally but globally much warmer late Eocene.
 |
| How to coexist when the global (and local) temperature rises? Lowlandia and Highlandia are two perfect NLRs with temperature-fitting altitudinal distribution and a highly informative modern-day coexistence space. To maintain coexistence, the ancient Lowlandia could have been growing at higher altitudes or the ancient Highlandia was more warmth-loving. When the general climate was warmer (or colder) than today, a parameter range shift, thus a violation of the actuo-palaeontological principle, is inevitable. |
Not matter how you turn it, the actuo-palaeontological principle is violated for at least one parameter, or even both. Make the math: if it was in general 2.5° warmer than today, and the critical taxa thrived at 500 m lower altitudes, Wu et al.'s CA result would be fitting reality while violating the basic principle for both parameters. If it would have been 2.5 °C colder than today in general, and critical taxa thrived higher up, it would be completely off reality. But still the CA protocol would ensure one gets a result.
I will not sketch once more how the authors should have done their analysis, since this is shown in great detail in our two 2016 papers. Just do it (or at least try).
What the vegetation really points to
With Wu et al.'s data, we get 'coexistence intervals' of 16.8–21.4 °C defined by Betula and Cedrus (either the value in Wu et al.'s table 1 or fig. 4 is a mistype), no outliers. And an altitude either within the expectation of the moist-adiabatic lapse rate, 1300–1600 m a.s.l. (Cedrus + Podocarpus; Abies, Artemisia, Ephedra, Picea, and Chenopodiaceae eliminated as too high), or one too high, 2000–2100 m (Ephedra, Picea too high; Lygodium, the temperate(!) fern, Castenea and Podocarpus too low); both intervals accommodate 4/5 of the NLRs. Today, at this altitude, we have subtropical conditions in the larger area (Cwa), and it was likely warmer in the late Eocene (we had vast forests covering e.g. the Arctic), which explains the "tropical-subtropical" in the abstract and conclusions.
[Wu et al. report and discuss 1300–2600 m as result (an upper boundary which according to their table 1 would be also too high for Carya, Castanea, Lygodium, Meliaceae and Podocarpus), so this is either a mistyper or some arcane analysis result.]
The most conspicuous observation is that typical, widespread, very common subtropical elements (especially in the Eocene) such as the infamous Engelhardioideae – the taxon defining M(A)AT of originally >17.5 °C (Mosbrugger & Utescher 1997), then >15.6 °C (all later studies), now >13.4 °C (Utescher et al. 2014; real modern MAT minimum tolerance lies at 10–11 °C: see Grimm & Denk 2012) are missing from the pollen taxon list. As are most other typical subtropical-tropical elements. In the late Eocene at mid-altitudes (1300–1600[–2600] m), with a global temperature about 5 °C warmer than today (cf. Zachos et al. 2001), they would need to be present.
The only taxa associated often associated with the subtropics (but not tropics) present are two conifers with a restricted, likely relict (i.e. unrepresentative) modern distribution: Keteleeria and "Taxodiaceae" (the latter using a long outdated classification concept [see even Wikipedia entry], another standard of the Coexistence Approach). The core part of the latter, the Taxodioideae, are known for forming coals being the main element of subtropical, low-altitude (sea-level) swamp forests (still today as relicts, see e.g. Earle 2011 onwards: the Japanese Cryptomeria; the Chinese Glyptostrobus; and U.S. Taxodium). But in the late Eocene, at sea-level at this latitude, the temperature must have been tropical. Other Cupressaceae genera that were included in the defunct Taxodiaceae have equally reduced modern distribution areas but appear to be distinctly temperate (some are even extending into the boreal zone) and without proper (SEM-based) pollen analysis it is impossible to decide which Cupressaceae Wu et al.'s "Taxodiaceae" actually represent (and even with, it may be difficult). The family itself is quite old, and likely lost a lot of lineages, with relatively few survivors today.
Lacking a lot of typical subtropical elements (or truly tropical ones) one would expect to find in the late Eocene, the flora boosts (subtropical-)temperate ones: we apparently had a conifer-dominated forest (Abies, Picea, Pinus) with few deciduous elements: the birch (Betula) and the walnut lineage, the Juglandoideae (Juglans, Carya), sister of the Engelhardioideae (see this GWoN post on the family), possibly montane evergreen elements, too (Quercus, also here the pollen would need to be re-studied using high-resolution SEM, to make any use of it for palaeo-vegetation/-climate reconstruction). The combination of Pinus, Betula and Juglans would be typical for the temperate broadleaved and mixed forest (TBMF) biome [Wikipedia] of the Northern Hemisphere, the lists includes even the typical fern found in some TBMF forests (Polypodium).
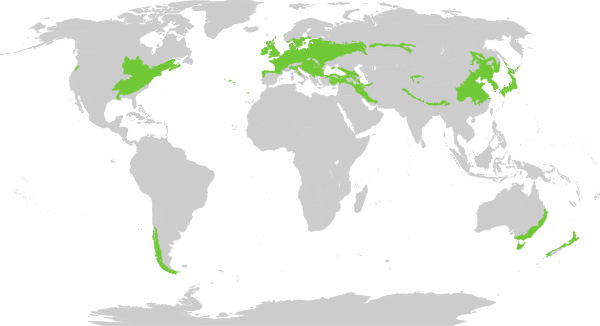 |
| The modern range of the temperate broadleaved and mixed forest biome, home to all taxa in Wu et al.'s list. Source: Wikimedia Commons, artwork by Terpsichores. |
Interesting are also the few other herbs: Artimisia and Chenopodioideae ("-aceae" in Wu et al.'s list, another obsolete family) are typical elements of cold steppe environments, e.g. the Artimisia steppes of continental Asia thriving under a BSk climate (as I recently learned, because of a paper on the Pikermian floras that finally is on the way to be published).
 |
| Found far away from seas in cold steppe (BSk) climates of the Northern Hemisphere: the Artimisia (sagebush) steppe (here: in an intramontane basin in Idaho, North America; source: Wikimedia Commons by Famartin; the accompanying article is a stub.) |
Most of the flora clearly doesn't point to the diffuse "tropical-subtropical, deciduous, coniferous, broadleaf forests" mentioned in the authors' abstract but a mosaic of montane temperate conifer-rich forests (taxon lists are typical for Cwb, less so Cfb climates, of the Northern Hemisphere) and (continental) steppe, similar to what can be found today in the region above 2500/3000 m a.s.l.
Assuming that the general temperature level in the late Eocene was about 5 °C higher than today and this would have been reflected in the local setting (Wu et al.'s transect is on the sea/southeast-facing side of the Qinghai-Tibetan Plateau), the flora – using the actuo-palaeontological principle and keeping the moist-adiabatic lapse rate in mind – indicates a palaeo-altitude of at least 3500/4000 m a.s.l.
Thus, providing further evidence that the Himalayan-related uplift of the Qinghai-Tibetan Plateau was already in full beauty by the late Eocene, as also depicted on the following map by C. R. Scotese that can be found on ResearchGate (published in 2013).
 |
| Right within the still growing mountains, not the worst place for a temperate mixed deciduous-conifer forest during a phase of ameliorated climate. |
Epilogue: The perils of confidential peer review
I want to conclude this post with two hypotheses that I have about the fact that CA-papers are still published despite fundamental flaws obvious to anyone with a little knowledge about vegetation and climate. They cannot be falsified or verified, because to do so, we would need to be able to lift the heavy curtain that currently clouds peer review: confidentiality.
Hypothesis 1. The authors were forced to add CA by the syndicate during confidential review
The CA analysis may have been a late add-on to the study, given the conciseness of the according chapters in Material & Methods and Results (p. 5):
4.2.2. The coexistence approach (CA)
Combining our results with the knowledge of the present-day distributions of fossil pollen NLRs, the diversity of the pollen taxa we observed would suggest the existence of a mixed deciduous, coniferous, broadleaf forest before ~36 Ma. Taking the pollen taxa as a whole into consideration, we used the CA to estimate a paleoelevation range of between 1300m and 2600m asl (Fig. 6 [maybe in an earlier manuscript version: current "Fig. 6" shows the "Correlations between δ13C and δ18O from primary (micrite) and secondary diagenetic (sparite) carbonates."]). Employing the CA, we deduced the region's mean annual air temperature (MAAT), arriving at a range of between 16.8 °C and 21.7 °C (Fig. 4). All paleovegetation, paleoelevation and paleoclimate data are listed in Table 1.
Combining our results with the knowledge of the present-day distributions of fossil pollen NLRs, the diversity of the pollen taxa we observed would suggest the existence of a mixed deciduous, coniferous, broadleaf forest before ~36 Ma. Taking the pollen taxa as a whole into consideration, we used the CA to estimate a paleoelevation range of between 1300m and 2600m asl (Fig. 6 [maybe in an earlier manuscript version: current "Fig. 6" shows the "Correlations between δ13C and δ18O from primary (micrite) and secondary diagenetic (sparite) carbonates."]). Employing the CA, we deduced the region's mean annual air temperature (MAAT), arriving at a range of between 16.8 °C and 21.7 °C (Fig. 4). All paleovegetation, paleoelevation and paleoclimate data are listed in Table 1.
In Material & Methods, they just cite several other Coexistence Approach papers that apparently reconstructed palaeo-altitudes in a similar way, including those of NECLIME's High Wizard and palaeoclimate expert Torsten Utescher, an editor's first-pick as peer when having a CA paper on the table not co-authored by him (NECLIME published two special issues in Palaeo^3, of course vetted by highest peer-review standards under the High Council's aegis, acting as guest editors).
Hypothesis 2: Coexistence Approach results published in Palaeo^3 (PPP) do not deserve the "peer reviewed" label
I'm still inclined to think that publishing and including illogical, erroneous palaeo-altitude and -temperature estimates was not the authors fault at all, but asked for during confidential review.
Authors were forced to include pseudo-scientific CA results by (more or less anonymous) peers in the past: NECLIMErs, in particular the High Wizard Utescher, a retired professor from Bonn (High Lord Mosbrugger will not have the time to do a lot of reviews being the director of the Senckenberg Society and Museum), may be recruited as (anonymous) peers. A natural editorial choice, after all they are well-published (HighWizard on ResearchGate; HighLord on ResearchGate; no GoogleScholar profiles, the incest would be too obvious, I suppose) thanks to Palaoe^3's pal review, true experts on palaeoclimate reconstruction (riding for 20 years the same dead horse). Veterans knowing their stuff. Or, to quote a relatively young NECLIME member: "It's unbelievable how much this man knows" (again, see the "anonymous" reviewer's comments to our 2012 paper). Result being a persistent demeaning of good (palaeobotanical) data and work that could be used to put forward meaningful qualitative or semi-quantitative results rather than pseudo-precise and -accurate quantitative house numbers.
But, under the line, just an example of many why we need basic rules for peer review documentation and transparency.
Regarding transparency, I like to point out that although Torsten Utescher advocates open data when it comes to the results of others (Utescher et al. 2014), he himself shows little interest in releasing the primary data he uses for the unexpected reconstruction of subtropical, monsoonal climates across the Eocene to Miocene of Eurasia (except Japan, they have apparently no good plant fossil lagerstätten there). Or why he keeps the used data in the closet when asked by a W.I.R.E.D journalist. Rather, a paper like the one of Tang et al. (2015) is retracted when exposed to the flashing light of transparent peer review (check out the Discussion tab).
Next time, dear Palaeo^3 editors (and CA-enforcing peer-pals), try a bit common sense, when assessing the scientific quality of the next Coexistence Approach paper to be published in Palaeo^3 or forcing unwitting authors to ride your dead horse during peer review.
Postscriptum If you want to read more funny papers using fuzzy logic, just use your institutional access (in case you have one — Palaeo^3 is a classic, exclusively public-funded, private-profit paywalled journal) to download Palaeo^3 papers by Torsten Utescher or any other main NECLIMEr, or any Coexistence Approach paper published in that journal by Chinese authors including a single non-Chinese co-author. But don't bother to contact the editors when depicting logical flaws, insane results, missing documentation etc. As the Editor-in-Chief pointed out to me: It's not their job to prevent pseudo-science being published in their journal and maintain basic standards of documentation and data access. They can only entertain formal comments. Subdued to the same, fine confidential peer review process than the original papers, of course.
Cited papers
Denk T, Grimm GW, Grímsson F, Zetter R. 2013. Evidence from "Köppen signatures" of fossil plant assemblages for effective heat transport of Gulf Stream to subarctic North Atlantic during Miocene cooling. Biogeosciences 10:7927–7942.
Grimm GW, Bouchal JM, Denk T, Potts AJ. 2016. Fables and foibles: a critical analysis of the Palaeoflora database and the Coexistence Approach for palaeoclimate reconstruction. Review of Palaeobotany and Palynology 233:216–235.
Grimm GW, Denk T. 2012. Reliability and resolution of the coexistence approach — A revalidation using modern-day data. Review of Palaeobotany and Palynology 172:33–47.
Grimm GW, Potts AJ. 2016. Fallacies and fantasies: the theoretical underpinnings of the Coexistence Approach for palaeoclimate reconstruction. Climates of the Past 12:611–622.
Maycock PF. 1994. The ecology of beech (Fagus grandifolia Ehrh.) forests of the deciduous forests of southeastern North America, and a comparison with the beech (Fagus crenata) forests of Japan In: Miyawaki A, Iwatsuki K, and Grandtner MM, eds. Vegetation in Eastern North America. Tokyo: University of Tokyo Press, p. 515 — data included in the online supplement archive to Grimm & Denk (2012)
Mosbrugger V, Utescher T. 1997. The coexistence approach -- a method for quantitative reconstructions of Tertiary terrestrial palaeoclimate data using plant fossils. Palaeogeography, Palaeoclimatology, Palaeoecology 134:61-86.
Tang H, Eronen JT, Kaakinen A, Utescher T, Ahrens B, Fortelius M. 2015. Strong winter monsoon wind causes surface cooling over India and China in the Late Miocene. Climates of the Past Discussions 11:63–93.
Thompson RS, Anderson KH, Bartlein PJ. 1999a. Atlas of relations between climatic parameters and distribution of important trees and shrubs in North America — Hardwoods. U S Geological Survey Professional Paper 1650–B:1–423.
Thompson RS, Anderson KH, Bartlein PJ. 1999b. Atlas of relations between climatic parameters and distributions of important trees and shrubs in North America — Introduction and Conifers. U S Geological Survey Professional Paper 1650–A:1–269.
Thompson RS, Anderson KH, Bartlein PJ. 2001. Atlas of relations between climatic parameters and distributions of important trees and shrubs in North America — Additional conifers, hardwoods, and monocots. U S Geological Survey Professional Paper 1650–C:1–386.
Thompson RS, Anderson KH, Pelltier RT, Strickland LE, Bartlein PJ, Shafer SL. 2012a. Quantitative estimation of climatic parameters from vegetation data in North America by the mutual climatic range technique. Quaternary Science Reviews 51:18–39.
Thompson RS, Anderson KH, Pelltier RT, Strickland LE, Shafer SL, Bartlein PJ. 2012b. Atlas of relations between climatic parameters and distributions of important trees and shrubs in North America—modern data for climatic estimation from vegetation inventories. U S Geological Survey Professional Paper 1650-F.
Thompson RS, Anderson KH, Strickland LE, Shafer SL, Pelltier RT, Bartlein PJ. 2006. Atlas of Relations Between Climatic Parameters and Distributions of Important Trees and Shrubs in North America—Alaska Species and Ecoregions. U S Geological Survey Professional Paper 1650-D:1-342.
Utescher T, Bruch AA, Erdei B, François I, Ivanov D, Jacques FMB, Kern AK, Liu Y-SC, Mosbrugger V, Spicer RA. 2014. The Coexistence Approach—Theoretical background and practical considerations of using plant fossils for climate quantification. Palaeogeography, Palaeoclimatology, Palaeoecology 410:58–73.
Wu J, Zhang K, Xu Y, Wang G, Garzione CN, Eiler J, Leloup PH, Sorrel P, Mahéo G. 2018. Paleoelevations in the Jianchuan Basin of the southeastern Tibetan Plateau based on stable isotope and pollen grain analyses. Palaeogeography, Palaeoclimatology, Palaeoecology DOI:10.1016/j.palaeo.2018.03.030.
Zachos JC, Pagani M, Sloan L, Thomas E, Billups K. 2001. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 292:686-693.
No comments:
Post a Comment
Enter your comment ...