Biodiversität, die Artenvielfalt, ist in aller Munde. Grundlage zum Messen der Biodiversität, ist fast immer (noch) die Anzahl der in einem Gebiet gefundenen Arten. Aber was ist eine Art? Ein paar Bildchen. Und Beispiele.
Durch unsere Arbeit an der coalface of evolution, kam ich recht früh mit den verschiedenen Artkonzepten in Kontakt. Besser gesagt, in Konflikt. Alle Artkonzepte (als wir danach suchten, fanden wir zwei Dutzend) waren theoretisch einleuchtend, aber in der Realität nicht allgemein anwendbar. Was für die eine Organismengruppe durchaus Sinn machte, funktionierte bei der nächsten nicht. Andere, wie das bei Zoologen immer noch sehr beliebte „biologische“ Artkonzept Ernst Mayrs, waren kaum überprüf- oder nachvollziehbar bei Pflanzen. Oder gar sinnfrei, wie das „kladistische“ Artkonzept.Die deutsche Wikipedia-Seite gibt einen recht netten Überblick sowohl über die konkurrierenden Artkonzepte, als auch über ihre Vor- und, nicht selten unüberwindlichen, Nachteile. Was fehlt, sind die passenden Bildchen.
Deswegen: Eine bebilderte Einleitung in die Wundervolle Welt der Arten (Fachbegriffe sind meist verlinkt mit der dt. Wikipedia; die Artikel sind meist ok, manche sehr kurz).
Über 300 Jahre alt, und immer noch hoch im Kurs: Linnés Art
Bis heute werden Arten in der Biologie mit Carl von Linnés „Binomen“ bezeichnet. Der Vorname, großgeschrieben, gibt dabei die Gattung an, der Nachname, kleingeschrieben, die Art. Meist ein beschreibendes Adjektiv, lateinisch oder latinisiert. Nach der systematischen Bibel, dem systematischen Code (ICN für Algen, Pflanzen und Pilze, ICZN für Tiere), sind alle „validen“, ordentlich nach den Regeln des Codes beschriebenen Gattungen, Untergattungen, Sektionen, Serien, Arten, Unterarten, Varietäten und Formen kursiv zu schreiben.
Zum Bespiel gehören alle natürlich vorkommenden Buchen in Zentral- und Westeuropa zur Art Fagus sylvatica L., der Rotbuche. Übersetzt: „Wald-Buche“. Ein Name der direkt von Linné stammt (daher das "L.") und Sinn macht: Buchen machen und stehen in Wäldern. Einige deutsche Buchenwälder sind sogar Teil des UNESCO Weltnaturerbes. Die Rotbuchen sind nicht die einzige Art in Europa: Bei seinen Reisen in den Kaukasus, fand Wolodymyr Lypskyi Buchen, die anders aussahen, und beschrieb sie als Fagus orientalis Lipsky, die „Orient-Buche“ (oder „Östliche Buche“). Greuter & Burdet entschieden 1981 (Med-Checklist Notulae 4, Willdenowia 11: 271-280), daß dies nur eine Unterart der Rotbuche ist: Heute reicht F. sylvatica subsp. orientalis (Lipsky) Greuter & Burdet vom südöstlichen Europa über die Türkei bis in den Iran. Dabei sehen sie anders aus, je nachdem wo man sie findet.
Das Aussehen eines Organismus, sein Phänotyp, bestimmt seine Art. Hier eine Sammlung von Individuen, die sich in ihrer Form, den Phänotypen, unterscheiden:

Der Artbeschreiber, der Taxonom, entscheidet dann, aus dem Bauch heraus (wie Greuter & Burdet) oder auf Grund seiner Beobachtungen in der Natur (berühmtes Beispiel: Darwins Finken), welches Merkmal „arttypisch“ ist und welches nicht: die „phänotypische“ oder „typographische“ Art ist geboren. In unserem Fall scheint es drei Arten zu geben: Fünfecke (Pentagone), Sterne, und Kreise.

Wie auch in der Natur, gibt es Übergangsformen. Seit nun schon Hunderten von Jahren, und bis heute, streiten Taxonomen darüber, wo man genau die schwarzen Linien zieht: die Artgrenzen. Sieht man genau hin: ein Teil der Kreise sind eigentlich Zahnräder, Sterne mit sehr vielen, sehr feinen Strahlen. Und wenn man die Kanten einknickt, werden Pentagone zu fünf-strahligen Sternen.

Warum gestritten wird? Entdeckt man eine neue Art (hier: die Zahnräder), wird man unsterblich: der eigene Namen bzw. das Autorenkürzel steht hinter der Art (bis jemand sie wieder abschafft).
 |
| Offiziell akzeptiert, der „Eigentliche (Ur-) Feld-Ahorn“ (PDF vom Paper, open access) |
Es gibt auch selbstlose Gründe: Wenn Rote Listen aufgestellt werden, werden die allgemein akzeptierten Arten (aufgelistet in Floren und Faunen) gezählt und berücksichtigt. Wenn ich also will, daß eine bestimmte Population geschüzt werden soll von zumeist komplett ahnungslosen und wenig-kompetenten Politikern (~2/3 des dt. Bundestags besteht aus Anwälten, reines Lernstudium, Berufspolitikern und Naturreichen, also hauptberuflichen Lobbyisten), mache ich aus ihr eine Art.
Das Ende der Willkürlichkeit: absolute Ähnlichkeit
Man kann die Unterschiede der Phänotypen auch vermessen:„morphometrische“ oder „phänetische“ Arten basieren auf demselben Ähnlichkeitsprinzip, aber auf (semi-)quantitativen, gemessenen Daten. Idealerweise von sehr vielen Individuen von verschiedenen Populationen die ein möglichst großes Gebiet und alle klassischen Arten einer Gattung abdecken.

Morphometrie, hier das Zählen der gut erkennbaren Strahlen, erlaubt die Aufteilung der polymorphen Art 2, den Sternen, in vier klar definierte Arten (2A–2D): 5-, 10-, 15- und 20-strahlige Sterne.
Unter der Annahme, daß die Strahligkeit Arten definiert, kann man Hybride (bzw. Hybridzonen, zwei Bsp.: Hausmaus und Krähe) postulieren. Die 25-zackigen Zahnräder (fast-Kreise) könnten Hybride von vielst-strahligen Sternen (Art 2D) und Kreisen (Art 3) sein. Die eingeknickten Pentagone hybride Übergangsformen zwischen den Pentagonen (Art 1) und den fünf-strahligen Sternen (Art 2A)
Bei Gattungen mit sehr hoher morphologischer Variabilität, wie z.B. den Eichen, wurden entsprechend viele Arten (>400 bei den Eichen: Oaks of the World) aber auch Hybride beschrieben (zig alleine in Europa, zwischen unterschiedlichsten Arten). Um typograpisch oder morphometrisch Hybriden zu erkennen, muß man erstmal die natürliche Variabilität bestehender Arten erfassen. Was sehr aufwendig ist. Hinzu kommt, daß die Natur ein Faible für Klinen hat: die Variabilität überlappt, statt ein klareres Bild ergibt sich ein diffuseres: der Übergang von Fünfecken zu 5-strahligen Sternen könnte solch eine Kline sein: Art 2A wäre dann nur eine Unterart von Art 1. Hier ein Beispiel basierend auf der Doktorarbeit meines langjährigen wissenschaftlichen Mittäters Thomas Denk (GoogleScholar), Steirischer Naturbursche und wissenschaftlicher Kurator am Naturhistorischen Museum in Stockholm.
 |
| Anzahl der Sekundärvenen (secondaries) und Blattmorphologien der Buchen von Spanien bis Iran. |
Für die Daten hinter dem Bild, besuchte Thomas sämtliche Populationen der Orient-Buchen in Bulgarien, Griechenland, der Türkei, Georgien und dem Iran, obwohl Greuter & Burdet sie schon als Unterart abgetan hatten. Und zahlreiche der noch ordinäreren Rotbuchen. Wissenschaftlich aufgearbeitet in fünf Publikationen in sogenannten low-impact Journals zwischen 1999 und 2004. Hoher Aufwand, unsicherer Profit (wir echten Wissenschaftler leben für und von Publikationen: bestenfalls Paper in high-impact Journals). Daher gibt es immer weniger, umfassende morphologisch-morphometrische Datensätze und Arbeiten.
Aber genetische.
Die Produktion genetischer Daten wird immer einfacher und leichter. Wir haben heute zahlreiche Möglichkeiten, den Genotyp eines Individuums oder einer Population zu erfassen. Genregionen, oder auch ganze Genome (das von SARS-CoV-2, dem Covid-19-Erreger, hat z.B. nur knapp über 30.000 Nukleotide; hier das Paper [open access] zum entsprechenden Genom-Browser) können sequenziert und verglichen werden. Der Vorteil: jeder kann genetische Daten auswerten, es ist kaum Vorwissen nötig. Die Computeralgorithmen ersetzen den Taxonom. Man legt dann einfach einen threshold fest, z.B. 5% Sequenzunterschied, und kriegt mit wenig Aufwand seine genetisch-definierten phänetischen Arten.
Bringen wir also ein wenig Farbe ins Spiel.

Da der Phänotyp nur bedingt vom Genotyp abhängt, kann es zu substanziellen Unterschieden kommen, was ähnlich und was unähnlich ist: die morphologische Art ↔ genetischen Art.
- Die Genotypen zeigen, daß die eingenickten Pentagone genetisch entweder sehr ähnlich den restlichen Pentagonen sind, zu Art 1 gehören, oder den wenig-strahligen Sternen: Art 2A.
- Es gibt es einen klaren Bruch zwischen den wenig- (Art 2A) und viel-strahligen Sternen (Art 2B): Morphologisch ist Art 2A variabler als Art 2B, genetisch sind sie gleichwertig.
- Außerdem finden wir zwei genetische Signaturen, die unser Artbild verändern bzw. durcheinanderbringen: eines der Pentagone (→ rote Art 1) ist genetisch ein Kreis (blaue Art 3A) und einer der Kreise ist genetisch einzigartig: neue hellblaue Art 3B.
Es gibt mehrere evolutive Prozesse, die zur Entkopplung von Phänotypen und Genotypen führen:
- Hybridisierung: Hybride können
- den Phänotyp eines Elter und den Genotyp des anderen Elter haben und weitergeben;
- durch Rückkreuzung mit nur einem Elter, fast ausschließlich den Geno- und Phänotyp dieses Elter übernommen haben;
- eine neuen Phänotyp zeigen, der durch die Kombinationen der Genotypen zustande kommt; oder ihre veränderte Expression;
- die phänotypischen und genotypischen Merkmale beider Elter mischen, wie bei dem blauen Fünfeck: der Genotyp ist der von Art 3, der Phänotyp der von Art 1.
Ein Paradebeispiel sind die Allee-Platanen (Platanus × acerifolia oder P. × hispanica), die „Ahornblättrigen“ bzw. „Spanischen Platanen“): sie sind hybriden Ursprungs (deswegen das „ד), entstanden durch die Hybridisierung von im 17./18. Jhdt. eingekreuzten, weil frost-tolerierenden östlichen nordamerikanischen Platanen – der „Platane des Westens“ Platanus occidentalis – in die in Südeuropa natürlich vorkommenden, aber frostempfindlichen Platanen des (östlichen) Mittelmeerraums: der „Platane des Ostens“ P. orientalis. Sie zeigen sämtliche möglichen Kombinationen. Und auch bei ihren wilden Verwandten geht es, vor allem genetisch, ähnlich chaotisch zu.
 |
| Platanen und ihre Genlinien. Gestrichelt: Pseudogene. Mehr dazu (freier Zugang, aber Englisch) in Grimm & Denk 2008, 2010, Denk et al. 2012, De Castro et al. 2013 |
- (Asymmetrische) Introgression, z.B. über früher herumfliegenden Pollen oder wenig wählerische Weibchen. Wie bei der Hybridisierung, kann Introgression zur Bildung intermediärer Phänotypen führen. Genauso häufig sieht man es aber den introgradierten Individuen und Populationen nicht an: die Introgression ist kryptisch (sog. ghost introgression – „Geister-Introgression“): ohne genetische Untersuchungen würde man nie auf die Idee kommen, das blaue Pentagon hätte irgendwas mit den Kreisen zu tun. Oder das Platanus palmeri, die südöstlichen Populationen der P. occidentalis, sich phänotypisch unterscheidet, weil sie halt auch mexikanische/kalifornische Eltern hatte.
Beispiele für kryptische, asymmetrische, und alte Introgressionen bzw. Hybridisierunen finden sich zu Hauf in den Buchengewächsen, den Fagaceen: ihre Plastome, die Genome der Chloroplasten, ausschließlich von den Müttern vererbt (wie bei vielen höheren Pflanzen), sind weitgehend entkoppelt sind von denen der Nukleome, der Genome des Zell-Kerns, vererbt von beiden Eltern, und den Phänotypen (für Details s. Oaks systematics and complete plastome trees). Nukleome und Phänotypen stimmen hingegen weitgehend überein, und häufig bis auf Artebene. Abgesehen natürlich von ein paar blauen Pentagonen.
- Detektions- und Sortierungsphänomene wie sample error [engl. Wikipedia] und incomplete lineage sorting [wieder nur engl.]: Sequenziert man mehr Genregionen, mehr Individuen in der Kontaktzone von Art 1 und Art 3A, oder gar die Gesamtgenome der Kreise und Pentagone, würde man vielleicht mehr blau finden, oder sogar rot in den Kreisen. Vielleicht ist der Blau-Anteil in den Pentagonen auch einfach verloren gegangen, während der Rot-Anteil in einigen Kreisen erhalten worden ist (das wäre eine Folge von incomplete lineage sorting). Auch hier sind die Platanen und Fagaceen wieder gute Beispiele.
Kryptische und pseudokryptische Arten
Einer der Kreise ist genetisch klar isoliert. Art 3B ist eine sogenannte „kryptische“ Art: Populationen und Individuen, die sich phänotypisch nicht von Art 3A unterscheiden lassen. Sehr häufig findet man kryptische Arten bei Organismen mit limitierten Morphologien (z.B. Bakterien, Einzellern, primitiven Pilzen, Tieren und Pflanzen). Aber auch bei gut-differenzierten Gruppen, deren Arten schon vor 100 Jahren beschrieben wurden. Die einzige rezente (heute lebende) Art, die mein Autorenkürzel trägt (Acer orthocampestre, s.o.), ist eine kryptische Ahorn-Art.
 |
| Kryptische Artbildung: Obwohl morphologisch ununterscheidbar, sind die kaukasischen Feldahorne (Acer orthocampestre) genetisch weiter weg von ihren westlichen Lookalikes (A. campestre s.str.) als die japanische, phänotypisch immer noch ähnliche Schwesterart (A. miyabei) und trotz der relativen geographischen Nähe. |
Bei den Buchen gibt es auch zwei Fälle von kryptischen Arten: Fagus hayatae (Taiwan) ↔ pashanica (C. China) und F. engleriana (weitverbreitet in China) ↔ F. multinervis (Ulleungdo, eine Vulkaninsel vor Korea).
 |
| Phylogenetischer Baum der Buchen basierend auf den Gendaten von Liang et al. (nach Cardoni et al. The Plant Journal 109: 909–926, 2021) Auffällig die phylogenetischen-genetischen Distanzen zwischen geographisch isolierten Populationen disjunkter Arten. Gleiche Farbe = gleiche Phänotypen (grau: Phänotyp unklar, da unbekannt ob von natürlichen Populationen oder aufgeforsteten). |
Erkennt man auf Grund der Genetik eine neue Art und stellt bei der Überprüfung der Phänotypen, der klassischen Arten, fest, daß man bisher einfach die Unterschiede unterschätzt, übersehen oder ignoriert hat, spricht man von einer „pseudokryptischen“ Art. Ein Beispiel sind die nun vier Arten der Giraffe, aber auch die noch nicht formalisierten 3–4 Arten der Orient-Buchen (im Baum oben ist eine davon abgedeckt) oder die mexikanischen Buchen (offiz. lt. Flora of North America: F. grandifolia; orange im Baum)
Die Quadratur des evolutionären Kreises durch Ernst Mayr
Als wohl größter Evolutionsbiologe seiner Zeit mit Kenntnissen der Populationsgenetik (und Feind von Hennig und dessen Kladistik, s. w. u.), entschied Mayr, daß nur solche Arten gute, natürliche Arten sind, deren Populationen sich nicht mehr vermischen können.
Das „biologische“ oder „populationsgenetische“ Artkonzept sagt: solange du reproduktionstechnisch nicht getrennt bist – deine Nachkommen fertil, fortpflanzungsfähig sind – gehörst du zur selben Art. Sind eure Nachkommen nicht lebens- oder fortpflanzungsfähig, haben sich eure Arten getrennt.
Wir schauen also welche Populationen das Kriterium erfüllen, direkt durch Kreuzungsexperimente, oder indirekt indem wir z.B. den Genfluß messen zwischen Populationen, und haben unsere „natürlichen“ biologischen Arten.

Alle Menschen, unabhängig ihrer Herkunft oder Phänotypen – Mayr selbst war, wie viele Zoologen seiner Zeit, ein Fan des biologischen Konzepts der Rasse, nicht zu verwechseln mit dem politisch-gesellschaftlichen Konzept (beide haben nur gemein, daß sie wissenschaftlich unhaltbar und sinnlos sind) – können fertile Nachkommen zeugen, wir sind also alle von der selben Art: Homo sapiens. Unsere Vorfahren, wenn sie auf Neandertaler trafen, hatten die Wahl zwischen Jagen+Braten und Paaren, und machten wohl beides. Bei heutigen Nicht-Afrikanern geht man von bis zu 5% Neandertaler-Anteil aus (schlechte Nachricht für alle Rassisten: die einzigen genetisch reinen modernen Menschen, sind die der Subsahara, simplifiziert: je hellhäutiger desto Höhle). Der Neandertaler ist also keine eigene Art (H. neanderthalensis), sondern, dem biologischen Artkonzept nach, nur eine Unterart des H. sapiens: H. sapiens neanderthalensis – wörtlich, der „weise [nicht weiße!] Mensch aus dem Neandertal“. Und je mehr Daten, darunter ancient DNA aus Knochenfunden, zusammenkommen, desto klarer wird: Kaum eine Art der Gattung Homo war reproduktionstechnisch komplett getrennt. Im Ursprung sind wir alle Bastarde.
 |
| Ein Stammbaum der Gattung Mensch (Homo): eine Artkoralle (Coral of Life). Quelle: Wikipedia Commons. |
Pferde (Equus Untergattung Equus) und Esel (Equus Untergattung Asinus) gehören zu verschiedenen Arten: Maultiere (E. africanus asinus♀ × E. ferus caballus♂), ihre Hybriden, sind unfruchtbar.
Hund und Wolf sind immer noch dieselbe Art (Canis lupus); Kojote (Canis latrans), Hund und Wolf eigentlich auch nach Mayrs biologischem Artkonzept, aber (in Reinform) phänotypisch und genotypisch unterschiedlich genug, daß die Zoologen sie weiterhin als zwei Arten sehen. Experten erkennen die Hybriden (Art-Kreuzungen) von Wolf/Hund und Kojote sofort, weil die Mutterarten klare phänetische Arten sind: sich im Phänotyp (und Genotyp) unterscheiden. Und trotz fehlender Reproduktionsbarriere – wie sie zwischen Hund und Rotfuchs, Vulpes vulpes, besteht: die unterschiedliche Chromosomensätze verhindern die Meiosis, die sexuelle Zellteilung. Hund/Wolf und Koyote sind phänetische-semi-biologische Arten, weil sie auf Grund ihres Verhaltens, Lebensweise und Lebensraums eher Inzucht (i. w. S., inbreeding) als Auskreuzung (outcrossing) betreiben.
In Zoos sind die vier neuen Giraffenarten kaum auseinanderzuhalten: sie sind nur genetisch und, bis zu einem gewissen Grad, phänotypisch in der Natur trennbar, weil die Individuen der schrumpfenden Wildpopulationen (vormaligen Unterarten) nicht mehr in Kontakt kommen können und Inzucht und genetische Verarmung (Generosion) fördern. Hätte der Mensch ihren Lebensraum nicht fragmentiert, könnten und würden sie sich kreuzen. Sie sind künstliche, anthropogene quasi-biologische Arten.
Eisbären, Ursus maritimus (der „Bär des Meeres“), und Grizzlies, eine Unterart der Braunbären (Ursus arctos horribilis; der „Schreckliche Arktis-Bär“)? Ursprünglich nur aus Zoos bekannt, haben die Kanadier begonnen jeden „Grolar-Bear“ zu erlegen, weil sie interfertil sind. In den Kontakt kommen die beiden phänotypisch sehr, und bis zu einem gewissen Grad, genotypisch unterschiedlichen Arten, weil wir den Lebensraum des Eisbären vernichten (2021 war wieder Rekordjahr für CO2-Ausstoß; 2022 wird den Rekord brechen).
Interfertilität und hemmungsfreies Ignorieren der Artgrenzen ist auch der Grund warum im Harz Hybrid-verdächtige süße Puschel eingefangen werden: es soll vermieden werden, daß der Genpool der Europäischen Wildkatzen (Felis silvestris – „Wald-Katze“) durch die ursprünglich aus dem Orient stammenden Hauskatzen (Felis catus – „Kater-Katze“) kontaminiert wird.
Dazu kommt das Phänomen der partiellen Infertilität, z.B. bei Ring-Arten (Ringspezies): (Unter-)Art A und B sind fertil, B und C auch, aber nicht A und C.
Und bei Pflanzen, Pilzen und allem anderen hat die Natur zahlreiche Wege gefunden, Artgrenzen oder genetische Inkompatibilität aufzuweichen: Asexuelle Fortpflanzung (z.B. Agamospermie), verschiedene Formen der Rekombination, Allopolyploidiserung usw.
In der Realität sieht es sehr häufig so aus wie im Schemabild: je mehr Daten ich generiere bzw. erfasse, desto einfacher wird es phänetische, auf Ähnlichkeit basierte Arten zu definieren, aber umso schwieriger wird es die Reproduktionsbarrieren, Mayrs Arten, zu identifizieren. Und selbst wenn ich es kann, sind die gefundenen Arten u.U. sehr schwer zu charakterisieren: warum sollte man gut-erkennbare Arten, wie z.B. die beiden häufig zusammen vorkommenden deutschen Eichenarten, Quercus robur (Stieleiche: lang-gestielte Früchte, Blattbasen mit „Öhrchen“) und Q. petraea (Traubeneiche: nicht oder kurz-gestielte Früchte, zulaufende Blattbasen), von Schulkindern (schon selbst getestet), Förstern und Botanikern unterscheidbar, aufgeben, nur weil sie noch nie von Mayr gehört haben und hier und da Genmaterial austauschen?
 |
| Relative genetisch-morphologische Kohärenz bei Stiel- und Traubeneichen im selben Wald. Von Neophytou, Forstarchiv 83, S. 34–40 (2012) |
Theorie und Praxis: Graphentheorie trifft Evolution
Im Gegensatz dazu hat das „kladistische“ Artkonzept, oft auch „phylogenetisches“ Artkonzept genannt, sehr klare und leicht zu ermittelnde Kriterien: zurückgehend auf die kladistische Systematik von Willi Hennig (der dabei einen Riesenbogen um Arten gemacht hat: nahezu jeder populationsgenetische Prozeß ist a-kladistisch), entspricht eine Art einer „monophyletischen“ – im Sinne Hennigs aber nicht Haeckels (Ashlock schlug daher 1971 vor es „holophyletisch“ zu nennen) – Gruppe von Individuen und/oder Populationen. Eine kladistische Art umfaßt alle Nachfahren des selben (hypothetischen) Ur-Ahns: des most-recent common ancestors. Im Falle des Menschen und oben gezeigtem Baum, gibt es nur 2 kladistisch valide Arten: Homo florensis und H. sapiens für alle anderen Homo (als erster beschriebenen Art).
Hennig hat seine Bäume noch handgemalt, basierend auf (seiner) Annahme, welches Merkmal Holophylie anzeigt (seine „Synapomorphien“). Heutzutage füttern wir einfach den Computer mit den vorhandenen (meist, und bestenfalls, genetischen) Daten und raus kommt ein phylogenetischer Baum. Der wird gewurzelt (meist mit Außengruppen), und wir können alle clades (dt. auch Klade), die (vermeintlich) holophyletischen Gruppen, einfach ablesen.
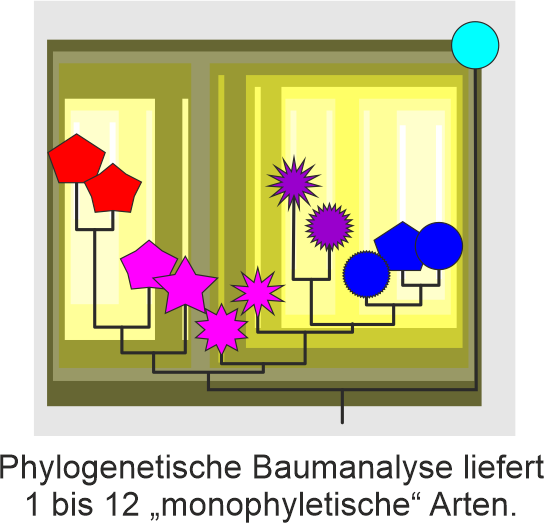
Arten definiert durch von Computeralgorithmen generierten phylogenetischen Bäumen. Absolut objektiv, einfach zu ermitteln und zu interpretieren, und, für sich genommen, sinnlos: jeder Knoten in einem gewurzelten phylogenetischen Baum entspricht einem most-recent common ancestor, dem exklusiven, hypothetischen allerletzten gemeinsamen Vorfahren. Jeder Ast definiert einen clade, ein Holophylum (in der Theorie).
Ein Baum mit n Tips (Blättern, im Fall von Artphylogenien Populationen bzw. Individuen) definiert also immer eine bis zu n Arten. Ohne Ähnlichkeiten zu erfassen (phänotypische und/oder genotypische), bestimmte Merkmale rauszupicken (wie schon Hennig es machte für alle Gruppen oberhalb der Art), die allen Mitgliedern eines clades gemein sind, habe ich kein Kriterium um zu sagen: ab diesem Ast, Knoten beginnt eine Art.
Aber sehr einfach anzuwenden zur Überprüfung klassischer, phänetischer, biologischer oder sonstiger Artkonzepte.

Wenn also von „monophyletischen“ Arten die Rede ist, sind in der Regel meist Linnésche Arten gemeint, die in dem jeweiligen molekularen Baum der Autoren eines Papers einen clade ergeben haben.
Und sollten sie es nicht getan haben, nennt man sie einfach „non-monophyletic“ – nicht-monophyletisch. Die Mühe neue, kladistisch-definierte Arten zu beschreiben (s. PhyloCode; die botanischen und zoologischen Codes verlangen eine eindeutige Diagnose), mit dem Risiko, daß sie im nächsten genetischen Baum keinen clade bilden und damit invalide werden, macht sich kaum einer.
Was man nicht auf Wikipedia lesen kann, oder in den allermeisten Lehrbüchern, geschweige denn im Bio-LK am Gymnasium lernt oder dem Bio-Studium an der Uni, ist, daß in der biologischen Realität, der coalface of evolution, ein clade in einem computergenerierten phylogenetischen Baum weder eine notwendige, noch eine hinreichende Bedingung für Holophylie ist. Egal wieviele Daten man (blind) kombiniert (Bsp.: Scientia-ex-machina: explicit biogeographic inferences and the phylogenomic age). Ein clade ist lediglich ein guter Hinweis auf (quasi-)Holophylie; im besten Fall, ein solides Argument.
Wir kamen daher auch nie in Versuchung das kladistische Artkonzept auch nur in Betracht zu ziehen. Bei unseren Forschungsobjekten (meist ganz ordinäre Bäume) gab es selten den immer gleichen, einen Baum. Und es gab offensichtliche Daten- und branching (Rekonstruktions-) Artefakte: false positives – falsche Positive. Scientia-ex-machina, die Wissenschaft aus der Maschine, funktioniert nur solange die Realität in der Natur den mathematischen Modellen der Computerprogramme (halbwegs) entspricht.
Was sie im Falle der Evolution und Artentstehung nicht (unbedingt) tut. Bzw. tat: die heutigen Arten sind eben nicht das Produkt eines sich immer in zwei Äste verzweigenden Baums (siehe Wikipediaseiten zu clade/Klade), sondern eines mehr oder weniger komplexen evolutionären Netzwerks, einer Artkoralle (Metaphors for evolutionary relationships; Morrison 2014). Nicht nur beim Menschen, auch bei unseren Buchen.
 |
| Evolution der Buchen bis hin zu den heutigen Arten. Die Kreuze stehen für sekundäre Kontakte und Vermischung zwischen Arten (oder innerhalb Arten oder Artkomplexen; um das zu sagen bräuchte man nicht nur ein universelles Artkonzept, sondern auch eine Zeitmaschine) |
Gehe zurück auf Los, und ziehe alle Artkonzepte ein
Die schwere Faßbarkeit der Art, ab wann spricht man von Art oder Unterart, die Unvereinbarkeit verschiedener Artkonzepte und die zahlreichen Probleme bei ihrer konkreten Anwendung, ist wohl auch der Grund, warum die frühen Evolutionsbiologen (vor Mayr und den Kladisten) sich nicht an der Definition irgendwelcher Artkonzepte abgemüht haben. Für sie waren Arten eben keine biologischen Grundeinheiten, sondern einfach die Stufe zwischen Ökotypen (oder Rassen bei den alten Zoologen)/ Varietäten (bei den Botanikern) und Gattungen. Systematisch-nützlich, biologisch erklärbar – auf Grund geographischen, ökologischen, Verhaltens-biologischen oder genetischen Reproduktionsbarrieren bzw. -hindernisse – wiedererkennbar, aber nicht universell definierbar. Konstrukte wie alle anderen in den systematischen Hierarchien, die dem Fluß der Zeit, und der Veränderung (Evolution) ausgesetzt sind.
 |
| Arten als Gruppen ± kohärenter Populationen (farbige Kreise = Genotypen) in Raum (geographical distance) und Zeit (time). Primär „phänetisch“, meist mehr „biologisch“ als „kladistisch“, aber klar phylogenetisch: alle ancestor-descendant Paare, die (tatsächlichen) Vorfahren und ihre Nachfahren, sind bekannt. Für jede heutige Art (mit * markiert), kann ich den, oder die, letzten gemeinsamen Vorfahren, bestimmen. Mehr dazu im Post Monophyletic species. |
Was heute eine gute, leicht erkennbare Art ist, also genotypisch und/oder phänotypisch gut charakterisierbare Gruppe von Populationen mit einem gemeinsamen Ursprung – common ancestry, abstammend von derselben oder denselben (Mehrzahl!) Ur-Population(en) (Bild oben) – kann in 1 Mio. Jahren schon eine Gattung mit zahlreichen, reproduktiv weitgehend getrennten, quasi-biologischen Arten sein. Was heute eine Gruppe von nahverwandten aber gut-sortierten Art-clades ist, kann durch Hybridisierung und Rückkreuzung wieder zu einer, möglicherweise polymorphen Art werden. Oder ein Artkomplex in dem unterschiedliche Gene/Genregionen unterschiedliche Genealogien, und andere clades, ergeben. Stirbt der Mensch aus, werden die vier jüngst getrennten Giraffenarten wohl wieder fusionieren in einen phänotypisch (Netzmuster) variablen, und genotypisch anfangs noch geographisch sortierten Artkomplex, der sich zunehmend homogenisiert und vermischt. Hunde, Koyoten und Schakale werden sich in den Zivilisationstrümmern treffen und paaren, Phänotypen werden verschwinden (hochgezüchtete Rassehunde), andere entstehen. U.u. neue, hybride Arten, mit so hohem Reproduktionserfolg, daß die heutigen Arten verdrängt werden oder in ihnen aufgehen. Vielleicht gibt es eine neue Bären(unter)art: den „Whizzly“, hellere Grizzlies in denen das genetische Erbe, die genetic legacy, der Eisbären in den dann vielleicht art-reicheren Braunbären weiterlebt. Entstanden nicht aus einem (dem most-recent common ancestor) oder einer Gruppe reproduktiv-isolierter Populationen, sondern einer ganzen Reihe von Grolar-Hybriden, die sich sowohl mit Grizzlies als auch den letzten Eisbären vergnügt hatten.
Praxis statt Theorie
Für uns paßte die Lösung von Mallet (A species definition for the Modern Synthesis. Trends in Ecology and Evolution 10: 294-299,1995 [PDF]): Art ist, was charakterisierbar (abgrenzbar) ist innerhalb von evolutionären (phylogenetischen) Linien. Empirisch definiert, datenbasiert. Was wiedererkennbar ist. Was einen systematischen Mehrwert hat. Verschwimmen die Arten bei Kontakt, sind es Unterarten. Gibt es keine Kontaktzone, liegt es immer noch am Experten, dem Feldbiologen/ Taxonom, ob Art oder Unterart.
Ob der gemeinsame Ursprung, der common origin, dabei „inclusive“ (holophyletisch, valide im kladistischen Sinn) oder „exclusive“ (paraphyletisch, invalide im kladistischen Sinn) ist, ob der Ursprung singulär (dichotom) oder komplex und retikulat ist, hat meist wenig Relevanz für das Bestimmen von Arten. Selbst die besten, solidesten Phylogenien, die rekonstruierten Bäume oder (besser:) Netzwerke, können nur den Rahmen vorgeben, für das finale Bild braucht es konkrete, vergleichbare Daten.
 |
| Empirische Arten Bsp. 1: Selbst kryptische Arten können trivial sein. Schon die simple Einfärbung der genetischen Distanzen zwischen den beprobten Individuen (grün: sehr ähnlich; rot: unähnlich) zeigt: die Feldahorne (Acer campestre) der Colchis (Kaukasus) sind schlicht anders. Sie stellen eine einzigartige genetische Resource dar, vergleichbar mit den schon bekannten anderen Arten der Sektion Platanoidea. |
 |
| Empirische Arten Bsp. 2: Artdiagnose im Zeitalter der Genetik. Der Abschnitt Genotaxonomy listet die Sequenz-Besonderheiten der neu-beschriebenen Art Drosanthemum nollothense hinsichtlich der Genregionen, die auch in den phylogenetischen Analysen benutzt wurden (Abschnitt Phylogeny). Kopiert aus Liede-Schumann et al. (2019, Bradleya 37: 226–239) |
Art oder nicht Art? Statt sich metaphysisch-konzeptionell zu fragen „Was ist eine Art?“, sollte man sich konkret fragen „Warum unterscheide ich diese Art?“
Z.B. um die vorhandene genetische und morphologische Diversität repräsentativ abzubilden, wie bei den Beispielen: Ahornen, Buchen und Platanen.
Es kann aber auch ganz unwissenschaftliche Gründe haben: Wenn ich durch das Beschreiben einer neuen, automatisch dann seltenen, Art die Plattmachung des nächsten Biotops für die …
- … Bestattung eines Bahnhofes und Schaffung exklusiven Wohnraums für Apex-Konsumenten …
- … einer Megafactory für unterm Strich Atom-, Fracking-Gas- bzw. Kohle-betriebene e-Individualpanzer …
- … generell für das Eisbär-tötende, aber Grolar-fördende Wachstum der Wirtschaft …
… verhindern kann, ist es eine gute Art
Und schnuppe ob primär „typographisch“, „phänetisch“, „biologisch“, „kladistisch“ oder datenbasiert empirisch.
No comments:
Post a Comment
Enter your comment ...